Oviraptor
| Oviraptor | |
|---|---|
 | |
| Espécime holótipo de Oviraptor | |
| Classificação científica | |
| Domínio: | Eukaryota |
| Reino: | Animalia |
| Filo: | Chordata |
| Clado: | Dinosauria |
| Clado: | Saurischia |
| Clado: | Theropoda |
| Família: | †Oviraptoridae |
| Subfamília: | †Oviraptorinae |
| Gênero: | †Oviraptor Osborn, 1924 |
| Espécie-tipo | |
| †Oviraptor philoceratops Osborn, 1924 | |
| Sinónimos | |
| |
Oviraptor (do latim "ladrão de ovos") é um gênero de dinossauro oviraptorídeo que viveu na Ásia durante o período Cretáceo Superior. Os primeiros restos mortais foram coletados da Formação Djadokhta da Mongólia em 1923 durante uma expedição paleontológica liderada por Roy Chapman Andrews, e no ano seguinte o gênero e a espécie-tipo Oviraptor philoceratops foram nomeados por Henry Fairfield Osborn. O nome do gênero se refere ao pensamento inicial de hábitos de roubo de ovos, e o nome específico pretendia reforçar essa visão, indicando uma preferência sobre ovos ceratopsianos. Apesar do fato de que vários espécimes foram referidos ao gênero, Oviraptor é conhecido apenas por um único esqueleto parcial considerado o holótipo, bem como um ninho de cerca de quinze ovos e vários pequenos fragmentos de um juvenil.
Oviraptor era um dinossauro emplumado bastante pequeno, estimado em 1,6–2 m de comprimento com um peso entre 33–40 kg. Ele tinha uma mandíbula inferior larga com um crânio que provavelmente tinha uma crista. As mandíbulas superior e inferior eram desdentadas e desenvolveram um bico córneo, que era usado durante a alimentação ao longo da morfologia robusta das mandíbulas inferiores. Os braços eram bem desenvolvidos e alongados, terminando em três dedos com garras curvas. Como outros oviraptorídeos, o Oviraptor tinha membros posteriores longos que tinham pés de quatro dedos, com o primeiro dedo reduzido. A cauda provavelmente não era muito alongada e terminava em um pigóstilo que sustentava penas grandes.
As relações iniciais do Oviraptor eram mal compreendidas na época e foi atribuído aos Ornithomimidae não relacionados pelo descritor original, Henry Osborn. No entanto, reexames feitos por Rinchen Barsbold provaram que Oviraptor era distinto o suficiente para justificar uma família separada, a Oviraptoridae. Quando descrito pela primeira vez, Oviraptor foi interpretado como um dinossauro ladrão e comedor de ovos, dada a associação próxima do holótipo com um ninho de dinossauro. No entanto, descobertas de vários oviraptorossauros em poses de nidificação demonstraram que este espécime estava na verdade chocando o ninho e não roubando nem se alimentando dos ovos. Além disso, a descoberta de restos de um pequeno juvenil ou filhote foi relatada em associação com o espécime do holótipo, apoiando ainda mais o cuidado parental.
História da descoberta

Os primeiros restos de Oviraptor foram descobertos em arenitos avermelhados da Formação Djadokhta do Cretáceo Superior da Mongólia, na localidade de Bayn Dzak (também conhecida como Penhascos Flamejantes), durante a Terceira expedição centro-asiática em 1923. Esta expedição foi liderada pelo naturalista estadunidense Roy Chapman Andrews e terminou na descoberta de três restos fósseis de terópodes novos para a ciência — incluindo os de Oviraptor. Estes foram formalmente descritos pelo paleontólogo norte-americano Henry Fairfield Osborn em 1924, que com base no novo material, nomeou os gêneros Oviraptor, Saurornithoides e Velociraptor. O gênero particular Oviraptor foi erguido com a espécie-tipo O. philoceratops com base no holótipo AMNH 6517, um indivíduo parcial sem a parte de trás do esqueleto, mas incluindo um crânio muito esmagado, vértebras cervicais e dorsais parciais, elementos peitorais incluindo a fúrcula com o braço esquerdo e mãos parciais, o ílio esquerdo e algumas costelas. Assim, este espécime foi encontrado deitado sobre um ninho de aproximadamente quinze ovos — um ninho que foi catalogado como AMNH 6508 — com o crânio separado dos ovos por apenas 10 cm de sedimento. Dada a proximidade de ambos os espécimes, Osborn interpretou Oviraptor como um dinossauro com hábitos de comer ovos, e explicou que o nome genérico, Oviraptor, é latim para "apanhador de ovos" ou "ladrão de ovos", devido à associação dos fósseis. O nome específico, philoceratops, é pretendido como "gosto por ovos de ceratopsianos", que também é dado como resultado do pensamento inicial do ninho pertencente a Protoceratops ou outro ceratopsiano. No entanto, Osborn sugeriu que o nome Oviraptor poderia refletir uma percepção incorreta deste dinossauro. Além disso, Osborn descobriu que Oviraptor era semelhante aos ornitomimídeos de corrida rápida não relacionados — na época, no entanto, considerados relacionados — com base nas mandíbulas sem dentes, e atribuiu Oviraptor aos Ornithomimidae.[1] Osborn havia relatado anteriormente o táxon como "Fenestrosaurus philoceratops",[2] mas isso foi desacreditado mais tarde.[1]

Em 1976, o paleontólogo mongol Rinchen Barsbold notou algumas inconsistências em relação à colocação taxonômica de Oviraptor e concluiu que esse táxon era bastante distinto dos ornitomimídeos com base em características anatômicas. Sob essa consideração, ele erigiu o Oviraptoridae para conter Oviraptor e parentes próximos.[3] Após a descrição inicial do gênero por Osborn, o ninho de ovos associado ao holótipo foi aceito como pertencente ao Protoceratops,[4][5] e os oviraptorídeos foram amplamente considerados como terópodes comedores de ovos.[6] No entanto, na década de 1990, a descoberta de vários espécimes de oviraptorídeos nidificantes e filhotes provou que Osborn estava correto em sua cautela quanto ao nome de Oviraptor. Essas descobertas mostraram que os oviraptorídeos chocavam e protegiam seus ninhos agachando-se sobre eles. Essa nova linha de evidências mostrou que o ninho associado ao holótipo de Oviraptor pertencia a ele e o espécime estava realmente chocando os ovos no momento da morte, não os caçando.[7][8][9]
Espécimes referidos

Após a nomeação de Oviraptoridae em 1976, Barsbold referiu seis espécimes adicionais para Oviraptor, incluindo dois espécimes específicos sob o número MPC-D 100/20 e 100/21.[3] Em 1986, Barsbold percebeu que os dois últimos não pertenciam ao gênero e, em vez disso, representavam um novo oviraptorídeo: Conchoraptor.[10] A maioria dos outros espécimes também provavelmente não pertencem ao próprio Oviraptor, e eles foram atribuídos a outros oviraptorídeos.[11][12] Um indivíduo parcial também com ovos da Formação Bayan Mandahu da Mongólia foi referido em 1996 por Dong Zhiming e Philip J. Currie, o espécime IVPP V9608.[9] No entanto, em 2010, Nicholas R. Longrich e os dois últimos paleontólogos expressaram suas incertezas em relação a esta referência, pois há várias diferenças anatômicas, como as proporções das falanges das mãos. Eles concluíram que este espécime era uma espécie diferente e indeterminada, não referenciável a este táxon.[13] Em 1981, Barsbold referiu o espécime MPC-D 100/42 ao Oviraptor, um indivíduo muito bem preservado e bastante completo da Formação Djadokhta.[14] Como os elementos conhecidos do Oviraptor eram tão fragmentários em comparação com outros membros, o MPC-D 100/42 se tornou a principal referência/representação deste táxon, sendo proeminentemente rotulado como Oviraptor philoceratops na literatura científica.[15][11]

Esta concepção foi refutada por James M. Clark e colegas em 2002, que notaram que este espécime de crista alta tem mais características do crânio em comum com Citipati do que com Oviraptor — que, de fato, não preserva uma crista — e pode representar uma segunda espécie do primeiro, ou um gênero inteiramente novo.[16] Em 1986, Barsbold descreveu uma segunda espécie de Oviraptor, "O. mongoliensis", com base no espécime MPC-D 100/32a que vem da Formação Nemegt.[10] No entanto, um reexame por Barsbold em 1997 encontrou diferenças suficientes neste espécime para nomear o novo gênero Rinchenia, mas ele não o descreveu com formalidade e este novo oviraptorídeo permaneceu como um nomen dubium.[17] Isso foi alterado pela paleontóloga polonesa Halszka Osmólska e equipe em 2004, nomeando formalmente o táxon Rinchenia mongoliensis.[18] O paleontólogo norte-americano Mark A. Norell e colegas em 2018 relataram um novo espécime de Oviraptor: AMNH 33092, que é composto de uma tíbia e dois metatarsos de um filhote ou juvenil muito pequeno. AMNH 33092 foi encontrado em associação com o holótipo e provavelmente fazia parte do ninho. Oviraptor agora é conhecido pelo holótipo com ovos associados e um juvenil/filhote.[19]
Descrição

O espécime do holótipo foi estimado em 1,6–2 m de comprimento com um peso variando de 33 a 40 kg.[20][21][22] Embora o holótipo não tenha em grande parte a região posterior do esqueleto, é provável que o Oviraptor tivesse dois membros posteriores bem desenvolvidos que terminavam em três dedos funcionais, sendo o primeiro vestigial, bem como uma cauda relativamente reduzida. Como evidenciado em oviraptorídeos relacionados, os braços eram cobertos por penas alongadas e a cauda terminava em um pigóstilo, que é conhecido por sustentar um leque de penas.[23][24]
Crânio
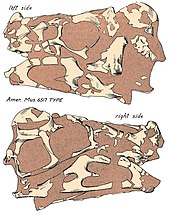
O crânio do Oviraptor era profundo e encurtado com grandes fenestras (aberturas) em comparação com outros dinossauros, e mede cerca de 17,9 cm de comprimento conforme preservado. O comprimento real pode ser maior, dado que o crânio do holótipo não possui várias regiões, como a pré-maxila. O crânio do holótipo não possui uma crista em quase sua totalidade, no entanto, as superfícies superiores dos ossos parietais e frontais fundidos indicam que provavelmente tinha uma crista bem desenvolvida, apoiada pelos ossos nasais e pré-maxilares (principalmente o último) do rostro. Oviraptor tinha uma maxila e dentário alongados, o que pode resultar em um focinho mais estendido em comparação com as mandíbulas altamente atarracadas de outros oviraptorídeos. O palato é rígido, estendido abaixo da linha da mandíbula e formado pelas pré-maxilas, vômeres e maxilas. Como em outros oviraptorídeos, pode ter tido um par de projeções semelhantes a dentes no palato que eram direcionadas para baixo. Como em outros oviraptorídeos, as narinas teriam sido relativamente pequenas e colocadas no alto do crânio. Oviraptor tinha mandíbulas desdentadas que terminavam em uma robusta ranfoteca (bico córneo) semelhante a um papagaio. A curvatura da ponta do dentário era voltada para baixo, mas menos pronunciada do que outros oviraptorídeos, como Citipati. Como um todo, a mandíbula inferior é um osso curto e profundo que cobre 19,5 cm.[16]
Esqueleto

Como na maioria dos membros de sua família, as espinhas neurais das vértebras cervicais do holótipo eram curtas, e os arcos neurais tinham formato de X. No entanto, as espinhas se tornam mais pronunciadas nas vértebras posteriores. As zigapófises das primeiras vértebras cervicais são configuradas paralelamente umas às outras, e as pós-zigapófises parecem não divergir significativamente da linha média, principalmente semelhantes a Citipati. As costelas cervicais são fundidas às vértebras no holótipo.[25][19] As espinhas neurais são retangulares na série anterior das vértebras dorsais quando vistas em uma vista lateral e maiores do que as espinhas das cervicais. Na vértebra dorsal mais anterior, várias pleuroceles (pequenos orifícios espaçados por ar) podem ser encontradas, que são semelhantes às de Khaan.[25]

A fúrcula de Oviraptor é muito distinta de outros oviraptorídeos por ter uma quilha da linha média na superfície anterior do hipocleídio - uma projeção direcionada para baixo no centro da fúrcula. Este osso tem formato de V, arredondado na seção transversal, preserva um hipocleídio alongado em forma de espícula, e o ângulo interclavicular é de cerca de 90°..[26] O escapulocoracóide é fundido no holótipo, no entanto, o coracoide está muito danificado. A escápula é ligeiramente arqueada e mede 23 cm de comprimento. Oviraptor tinha um braço relativamente alongado composto pelo úmero, rádio, ulna e manus.[1][25]
A fórmula falangeana do Oviraptor era 2-3-4, como visto na maioria dos outros terópodes e oviraptorídeos. A mão do Oviraptor tinha três dedos finos e semelhantes a pássaros, com cada dedo terminando em ungueais (osso da garra) achatados e recurvados de um lado para o outro. Ao contrário de alguns oviraptorídeos, o Oviraptor não sofreu redução do segundo e terceiro dedos em relação ao primeiro.[1][25][27] O considerado juvenil Oviraptor AMNH 33092 preserva material do membro posterior, compreendendo uma tíbia direita com metatarsos III e IV. Sua tíbia tem 5,87 cm de comprimento, indicando um indivíduo substancialmente menor do que o holótipo.[19] O ninho AMNH 6508 preserva ovos com cada tendo 14 cm de comprimento (alguns estão incompletos). No entanto, existe a possibilidade de que o esmagamento tafonômico possa tê-los comprimido em até 2 cm.[7][28]
Classificação
Oviraptor foi originalmente aliado aos ornitomimídeos por Osborn devido ao seu bico sem dentes. Osborn também encontrou semelhanças com Chirostenotes, que ainda é considerado um parente próximo de Oviraptor.[1] Em 1976, Barsbold erigiu uma nova família para conter Oviraptor e seus parentes próximos, tornando Oviraptor o gênero tipo dos Oviraptoridae.[3] Durante a redescrição do crânio do holótipo em 2002 por Clark e colegas, eles notaram que Oviraptor tinha uma maxila e dentário relativamente alongados. Essas características são menos pronunciadas em oviraptorídeos derivados e sugerem que Oviraptor pertence à base próxima dos Oviraptoridae.[16]
O cladograma abaixo segue uma análise de Gregory F. Funston e colegas em 2020:[29]
| Oviraptoridae |
| ||||||||||||||||||||||||
Paleobiologia
Alimentação

Quando descrito pela primeira vez em 1924 por Osborn, o Oviraptor foi originalmente presumido como comedor de ovos com base na associação do holótipo com um ninho que se pensava pertencer ao Protoceratops.[1] Em 1977, Barsbold propôs uma hipótese de mandíbula esmagadora. Ele argumentou que a força das mandíbulas inferiores robustas e provavelmente da ranfoteca (bico córneo) era forte o suficiente para quebrar as conchas de moluscos como amêijoas, que são encontradas na mesma formação geológica que o Oviraptor. Esses ossos fazem parte do osso principal da mandíbula superior ou maxila, que convergem no meio para formar um par de pontas. A ranfoteca e as mandíbulas inferiores, juntamente com a extensão de vários ossos do palato, teriam feito uma ferramenta de perfuração. Barsbold também sugeriu que os oviraptorídeos poderiam ter tido um estilo de vida semiaquático baseado na dieta baseada em moluscos, na alta localização das cavidades nasais, na musculatura aumentada da cauda e no tamanho maior do primeiro dígito manual.[30] Em um resumo de conferência de 1990, David K. Smith apresentou uma reavaliação osteológica do Oviraptor, onde rejeitou as declarações feitas por Barsbold. Ele não encontrou nenhuma evidência indicando um membro anterior especializado em locomoção aquática, e as mandíbulas, em vez de preservar um mecanismo de esmagamento, preservam superfícies de cisalhamento. Como o crânio é desdentado, de constituição leve e carece de várias áreas de inserção muscular fortes, Smith sugeriu que as folhas podem ter sido uma parte importante na dieta do Oviraptor.[31] No entanto, em 1995, Norell e colegas relataram os restos fragmentados de um lagarto na cavidade corporal do espécime holótipo, sugerindo que o Oviraptor era parcialmente carnívoro.[32]

Em 2008, Stig Olav K. Jansen comparou o crânio de várias espécies de oviraptorídeos aos de pássaros e tartarugas para investigar quais propriedades podem prever uma ranfoteca. Ele descobriu que as mandíbulas inferiores dos oviraptorídeos eram muito semelhantes às dos papagaios, e as mandíbulas superiores eram mais semelhantes às das tartarugas. Com base nessas observações, Jansen sugeriu que os oviraptorídeos eram onívoros, pois a ranfoteca fortemente desenvolvida, juntamente com os membros anteriores proeminentes, teria sido adaptada para capturar e rasgar pequenas presas. Além disso, as projeções pontiagudas do palato teriam contribuído para segurar a presa. Jansen apontou que uma dieta totalmente herbívora em oviraptorídeos parece improvável, pois eles não tinham tomia plana e larga (bordas cortantes das mandíbulas) para mastigar e eram incapazes de mover as mandíbulas inferiores para os lados. No entanto, ele considerou as mandíbulas inferiores fortes o suficiente para ter pelo menos elementos triturados como ovos, nozes ou outras sementes duras.[33]
Longrich e colegas em 2010 também rejeitaram uma hipótese durófaga (animais que praticam esmagamento de conchas), dado que tais animais normalmente desenvolvem dentes com superfícies de esmagamento largas. O formato pontiagudo dos ossos dentários nas mandíbulas inferiores sugere que os oviraptorídeos tinham uma ranfoteca de bordas afiadas usada para tosquiar alimentos. A região sínfise (união óssea) na frente do dentário pode ter dado alguma habilidade para esmagamento, mas como esta era uma área relativamente pequena, provavelmente não era a função principal das mandíbulas. Outro argumento contra eles terem sido comedores de moluscos é o fato de que a maioria dos oviraptorídeos foi encontrada em sedimentos que são interpretados como representando ambientes principalmente áridos ou semiáridos, como Oviraptor na Formação Djadokhta. A equipe também descobriu que oviraptorídeos e dicinodontes compartilham características cranianas como mandíbulas curtas, profundas e desdentadas; sínfises dentárias alongadas; aberturas mandibulares alongadas; e um palato pontudo. Animais modernos com mandíbulas que se assemelham às dos oviraptorídeos incluem papagaios e testudinídeos; o último grupo também tem projeções semelhantes a dentes na região palatina. Longrich e colegas concluíram que, devido às semelhanças entre oviraptorídeos e animais herbívoros, a maior parte de sua dieta provavelmente teria sido formada por material vegetal. As mandíbulas dos oviraptorídeos podem ter sido especializadas para processar alimentos, como vegetação xerófita - uma vegetação adaptada a ambientes com pouca água - que teria crescido em seus ambientes áridos, mas isso não é possível demonstrar, pois pouco se sabe sobre a paleoflora do Deserto de Gobi.[13]
Em 2018, no entanto, Funston e colegas apoiaram a hipótese da mandíbula esmagadora. Eles apontaram que o rostro atarracado e as mandíbulas inferiores robustas dos oviraptorídeos sugerem, de fato, uma mordida forte e cortante, que é bastante semelhante à dos papagaios. Funston e colegas consideraram que essas características anatômicas dos oviraptorídeos eram consistentes com uma dieta frugívora que incorporava nozes e sementes.[34]
Reprodução


Desde a descrição do espécime embrionário de Citipati em 1994, os oviraptorídeos se tornaram mais compreendidos: em vez de serem animais comedores de ovos, eles na verdade chocavam e cuidavam dos ninhos. Este espécime mostrou que o holótipo de Oviraptor era provavelmente um indivíduo sexualmente maduro que pereceu incubando o ninho associado com ovos.[7] Este novo comportamento em oviraptorídeos se tornou mais claro com o relato e breve descrição de um espécime adulto de nidificação de Citipati em 1995 por Norell e colegas. O espécime foi encontrado em cima de ninhadas de ovos, com seus membros posteriores agachados simetricamente em cada lado do ninho e os membros anteriores cobrindo o perímetro do mesmo. Esta postura de choca é encontrada hoje apenas em dinossauros aviários modernos e apoia uma ligação comportamental entre o último grupo e dinossauros não aviários.[8] Em 1996, Dong e Currie descreveram um novo espécime de oviraptorídeo nidificante da Formação Bayan Mandahu. Foi encontrado deitado em cima de um ninho composto por aproximadamente 6 ovos conforme preservados, e estes foram colocados em uma estrutura em forma de monte com um padrão circular. Como o espécime foi encontrado sobre o ninho com seus membros anteriores cobrindo os ovos e o pé traseiro parcialmente preservado perto do centro do ninho, Dong e Currie sugeriram que ele foi capturado e enterrado por uma tempestade de areia durante a incubação. Eles descartaram a possibilidade de oviraptorídeos serem ladrões de ovos, pois eles teriam consumido ou abandonado instintivamente o ninho muito antes de ser enterrado por uma tempestade de areia ou outro fenômeno meteorológico.[9]
Em 1999, Clark e equipe descreveram em detalhes o espécime de nidificação Citipati relatado anteriormente e discutiram brevemente o espécime holótipo de Oviraptor e sua associação com o ninho AMNH 6508. Eles apontaram que a posição exata em que o holótipo foi encontrado sobre o ninho não é clara, pois eles foram separados durante a preparação, e o ninho parece não estar totalmente completo com cerca de 15 ovos preservados, dos quais dois danificados. Além disso, a disposição semicircular do ninho indica que os ovos eram postos aos pares e em pelo menos três anéis, e este ninho era originalmente circular, semelhante a um monte.[28]
Thomas P. Hopp e Mark J. Orsen analisaram em 2004 o comportamento de incubação de espécies de dinossauros extintas e existentes, incluindo oviraptorídeos, para avaliar a razão do alongamento e desenvolvimento das penas das asas e da cauda. Dado que o espécime de nidificação de oviraptorídeo mais completo — na época, o espécime de nidificação Citipati de 1995 — foi encontrado em uma postura muito parecida com a de uma ave, com os membros anteriores em uma postura quase dobrada e a região peitoral, barriga e pés em contato com os ovos, Hopp e Orsen indicaram que longas penas e uma plumagem provavelmente estavam presentes na vida. As "asas" e a cauda dos oviraptorídeos teriam garantido proteção para os ovos e filhotes contra fatores climáticos como luz solar, vento e chuvas. No entanto, os braços deste espécime não eram extremamente dobrados como em algumas aves modernas, em vez disso, eles são mais estendidos, lembrando o estilo de grandes aves não voadoras, como o avestruz. A posição estendida do braço também é semelhante ao comportamento de chocar desta ave, que é conhecida por nidificar em grandes ninhadas como os oviraptorídeos. Com base na posição dos membros anteriores dos oviraptorídeos em nidificação, Hopp e Orsen propuseram a chocar como a razão ancestral por trás do alongamento das penas das asas e da cauda, pois havia uma necessidade maior de fornecer proteção ideal para ovos e juvenis.[35]

Em 2005, Tamaki Sato e equipe relataram um espécime incomum de oviraptorídeo da Formação Nanxiong. Este novo espécime foi encontrado preservando principalmente a região pélvica com dois ovos dentro e, portanto, indicando uma fêmea. O tamanho e a posição dos ovos sugerem que os oviraptorídeos mantiveram dois ovidutos funcionais, mas reduziram o número de ovos ovulados para um por oviduto.[36] David J. Varricchio e colegas em 2008 descobriram que o tamanho relativamente grande da ninhada de ovos de oviraptorídeos e troodontídeos é mais semelhante ao de pássaros modernos que praticam acasalamento polígamo e cuidados parentais masculinos extensivos, como aves ratitas, sugerindo hábitos semelhantes. Este sistema reprodutivo é mais provável que represente a condição ancestral para pássaros modernos, com o cuidado biparental (onde ambos os pais participam) sendo um desenvolvimento posterior.[37] Em 2014, W. Scott Persons e colegas sugeriram que os oviraptorossauros eram secundariamente incapazes de voar e várias das características em suas caudas podem indicar uma propensão para comportamento de exibição, como cortejo sexual. A cauda de vários oviraptorossauros e oviraptorídeos terminava em pigóstilos, uma estrutura óssea na ponta da cauda que, pelo menos em pássaros modernos, é usada para sustentar um leque de penas. Além disso, a cauda era notavelmente musculosa e tinha uma flexibilidade pronunciada, o que pode ter auxiliado nos movimentos de cortejo.[24]

Em 2018, Tzu-Ruei Yang e colegas identificaram camadas de cutícula em várias cascas de ovos de dinossauros maniraptoranos, incluindo aquelas de oviraptorídeos. Essas camadas específicas são compostas de proteínas, polissacarídeos e pigmentos, mas principalmente de lipídios e hidroxiapatita. Em pássaros modernos, elas servem para proteger os ovos da desidratação e invasão de microrganismos. Como a maioria dos espécimes de oviraptorídeos foram encontrados em formações de sedimentação baseadas em caliche, Yang e colegas sugeriram que os ovos revestidos de cutícula teriam sido uma estratégia reprodutiva adaptada para aumentar seu sucesso de eclosão em climas e ambientes tão áridos.[38]
Em 2019, Yang e colegas reavaliaram a hipótese de incubação de contato termorreguladora usando ninhos completos de oviraptorídeos da Formação Nanxiong e forneceram uma reconstrução detalhada da arquitetura da ninhada de oviraptorídeos. Eles notaram que espécimes adultos de oviraptorídeos em associação com o ninho não estavam necessariamente incubando os ovos, pois poderiam representar uma fêmea no processo de postura de ovos, e a ninhada de vários anéis impedia a transferência de calor suficiente do progenitor para os anéis internos dos ovos. Um ninho médio de oviraptorídeos foi construído como um monte suavemente inclinado com uma arquitetura altamente organizada: os ovos eram provavelmente pigmentados e dispostos em pares, com cada par disposto em três a quatro anéis elípticos. Como o progenitor provavelmente estava operando a partir do centro do ninho, esta região estava desprovida de ovos. Yang e colegas concluíram que o estilo de nidificação dos oviraptorídeos era tão único que carecem de análogos modernos; portanto, usar a reprodução dos oviraptorídeos pode não ser o melhor exemplo para informar sobre a evolução das estratégias reprodutivas das aves. Entretanto, a equipe não conseguiu determinar se o Oviraptor AMNH 33092 juvenil havia eclodido do ninho associado ao holótipo.[39]
Paleoambiente

Oviraptor é conhecido da localidade de Bayn Dzak da Formação Djadokhta na Mongólia, uma formação que remonta ao Cretáceo Superior, cerca de 71 milhões a 75 milhões de anos atrás.[40] O paleoambiente da Formação Djadokhta é interpretado como tendo um clima semiárido, com dunas de areia e cenários aluviais semelhantes ao moderno Deserto de Gobi. A paisagem de estepe semiárida era drenada por riachos intermitentes e às vezes era afetada por poeira e tempestades de areia, e a umidade era sazonal.[41][42] Embora esta formação seja amplamente considerada como um ambiente altamente árido preservado, vários corpos d'água de curta duração foram relatados na localidade de Ukhaa Tolgod, com base na sedimentação fluvial.[43] Além disso, acredita-se que mais tarde nos estágios Campaniano e no Maastrichtiano, o clima mudaria para o ambiente fluvial mais úmido visto na Formação Nemegt.[42]
A Formação Djadokhta é separada em um Membro Bayn Dzak inferior e um Membro Turgrugyin superior. Os restos conhecidos de Oviraptor foram produzidos pelo membro Bayn Dzak, que também produziu os dinossauros Bainoceratops, Pinacosaurus, Protoceratops, Saurornithoides, Velociraptor,[40] e Halszkaraptor.[44] Outra fauna de dinossauros deste membro inclui a da localidade Ukhaa Tolgod, composta por Apsaravis, Byronosaurus, Citipati, Gobipteryx, Khaan, Khol, Shuuvuia, Tsaagan,[43] e Minotaurasaurus.[45]
Ver também
Notas
- Este artigo foi inicialmente traduzido, total ou parcialmente, do artigo da Wikipédia em inglês cujo título é «Oviraptor», especificamente desta versão.
Referências
- ↑ a b c d e f Osborn, H. F. (1924). «Three new Theropoda, Protoceratops zone, central Mongolia». American Museum Novitates (144): 1−12. OCLC 40272928. hdl:2246/3223

- ↑ Osborn, H. F. (1924). «The discovery of an unknown continent». Natural History. 24 (2): 133−149
- ↑ a b c Barsbold, R. (1976). «О новом позднемеловом семействе мелких теропод Oviraptoridae fam. nov Монголии». Doklady Akademii Nauk SSSR (em russo). 226 (3): 685−688
- ↑ Coombs, W. P. (1989). «Modern analogs for dinosaur nesting and parental behavior». In: Farlow, J. O. Paleobiology of the dinosaurs. Col: Geological Society of America Special Papers. Geological Society of America Special Paper 238. Colorado: Boulder. pp. 21−54. ISBN 0-8137-2238-1. doi:10.1130/SPE238-p21
- ↑ Thulborn, R. A. (1992). «Nest of the dinosaur Protoceratops». Lethaia. 25 (2): 145−149. Bibcode:1992Letha..25..145T. doi:10.1111/j.1502-3931.1992.tb01379.x
- ↑ Currie, P. J.; Godfrey, S. J.; Nessov, L. A. (1993). «New caenagnathid (Dinosauria: Theropoda) specimens from the Upper Cretaceous of North America and Asia». Canadian Journal of Earth Sciences. 30 (10): 2255−2272. Bibcode:1993CaJES..30.2255C. doi:10.1139/e93-196
- ↑ a b c Norell, M. A.; Clark, J. M.; Dashzeveg, D.; Barsbold, R.; Chiappe, L. M.; Davidson, A. R.; McKenna, M. C.; Altangerel, P.; Novacek, M. J. (1994). «A theropod dinosaur embryo and the affinities of the Flaming Cliffs Dinosaur eggs». Science. 266 (5186): 779−782. Bibcode:1994Sci...266..779N. JSTOR 2885545. PMID 17730398. doi:10.1126/science.266.5186.779
- ↑ a b Norell, M. A.; Clark, J. M.; Chiappe, L. M.; Dashzeveg, D. (1995). «A nesting dinosaur». Nature. 378 (6559): 774−776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0
- ↑ a b c Dong, Z.; Currie, P. J. (1996). «On the discovery of an oviraptorid skeleton on a nest of eggs at Bayan Mandahu, Inner Mongolia, People's Republic of China». Canadian Journal of Earth Sciences. 33 (4): 631−636. Bibcode:1996CaJES..33..631D. doi:10.1139/e96-046
- ↑ a b Barsbold, R. (1986). «Хищные динозавры овирапторы» [Carnivorous dinosaurs oviraptors]. In: Vorobyeva, E. I. Herpetological research in the Mongolian People's Republic (em Russian). Institute of Evolutionary Morphology and Ecology of Animals: Doklady Akademii Nauk SSSR. pp. 210–223 !CS1 manut: Língua não reconhecida (link)
- ↑ a b Maryańska, T.; Osmólska, H.; Wolsan, M. (2002). «Avialan status for Oviraptorosauria» (PDF). Acta Palaeontologica Polonica. 47 (1): 97−116
- ↑ Kundrát, M.; Janáček, J. (2007). «Cranial pneumatization and auditory perceptions of the oviraptorid dinosaur Conchoraptor gracilis (Theropoda, Maniraptora) from the Late Cretaceous of Mongolia». Naturwissenschaften. 94 (9): 769−778. Bibcode:2007NW.....94..769R. PMID 17530209. doi:10.1007/s00114-007-0258-7
- ↑ a b Longrich, N. R.; Currie, P. J.; Dong, Z. (2010). «A new oviraptorid (Dinosauria: Theropoda) from the Upper Cretaceous of Bayan Mandahu, Inner Mongolia». Palaeontology. 53 (5): 945−960. Bibcode:2010Palgy..53..945L. doi:10.1111/j.1475-4983.2010.00968.x
- ↑ Barsbold, R. (1981). «Беззубые хищные динозавры Монголии» [Toothless carnivorous dinosaurs of Mongolia]. Transactions, Joint Soviet–Mongolian Palaeontological Expedition (em Russian). 15: 28−39 !CS1 manut: Língua não reconhecida (link) Periódico traduzido
- ↑ Barsbold, R.; Maryańska, T.; Osmólska, H. (1990). «Oviraptorosauria». In: Weishampel, D. B.; Osmolska, H.; Dodson, P. The Dinosauria 1st ed. Berkeley: University of California Press. pp. 249−258. ISBN 9780520067271
- ↑ a b c Clark, J. M.; Norell, M. A.; Rowe, T. (2002). «Cranial Anatomy of Citipati osmolskae (Theropoda, Oviraptorosauria), and a Reinterpretation of the Holotype of Oviraptor philoceratops» (PDF). American Museum Novitates (3364): 1−24. doi:10.1206/0003-0082(2002)364<0001:CAOCOT>2.0.CO;2. hdl:2246/2853
- ↑ Barsbold, 1997. Oviraptorosauria. In Currie and Padian (eds.). Encyclopedia of Dinosaurs. 505-509.
- ↑ Osmólska, H.; Currie, P. J.; Barsbold, R. (2004). «Oviraptorosauria». In: Weishampel, D. B.; Dodson, P.; Osmólska, H. The Dinosauria 2nd ed. Berkeley: University of California Press. pp. 165−183. ISBN 0520242092
- ↑ a b c Norell, M. A.; Balanoff, A. M.; Barta, D. E.; Erickson, G. M. (2018). «A second specimen of Citipati osmolskae associated with a nest of eggs from Ukhaa Tolgod, Omnogov Aimag, Mongolia». American Museum Novitates (3899): 1−44. doi:10.1206/3899.1. hdl:2246/6858
- ↑ Werner, J.; Griebeler, E. M. (2013). «New Insights into Non-Avian Dinosaur Reproduction and Their Evolutionary and Ecological Implications: Linking Fossil Evidence to Allometries of Extant Close Relatives». PLOS ONE. 8 (8): e72862. Bibcode:2013PLoSO...872862W. PMC 3749170. PMID 23991160. doi:10.1371/journal.pone.0072862
- ↑ Paul, G. S. (2016). The Princeton Field Guide to Dinosaurs 2nd ed. Princeton, New Jersey: Princeton University Press. 178 páginas. ISBN 9780691167664
- ↑ Campione, N. E.; Evans, D. C. (2020). «The accuracy and precision of body mass estimation in non-avian dinosaurs». Biological Reviews. 95 (6): 1759–1797. PMID 32869488. doi:10.1111/brv.12638
- ↑ Fanti, F.; Currie, P. J.; Badamgarav, D.; Lalueza-Fox, C. (2012). «New specimens of Nemegtomaia from the Baruungoyot and Nemegt Formations (Late Cretaceous) of Mongolia». PLOS ONE. 7 (2): e31330. Bibcode:2012PLoSO...731330F. PMC 3275628. PMID 22347465. doi:10.1371/journal.pone.0031330
- ↑ a b Persons, W. S.; Currie, P. J.; Norell, M. A. (2014). «Oviraptorosaur tail forms and functions». Acta Palaeontologica Polonica. doi:10.4202/app.2012.0093
- ↑ a b c d Balanoff, A. M.; Norell, M. A. (2012). «Osteology of Khaan mckennai (Oviraptorosauria, Theropoda)». Bulletin of the American Museum of Natural History. 372 (372): 1−77. OCLC 884738416. doi:10.1206/803.1. hdl:2246/6413
- ↑ Nesbitt, S. J.; Turner, A. H.; Spaulding, M.; Conrad, J. L.; Norell, M. A. (2009). «The theropod furcula». Journal of Morphology. 270 (7): 856−879. PMID 19206153. doi:10.1002/jmor.10724
- ↑ Lü, J.; Currie, P. J.; Xu, L.; Zhang, X.; Pu, H.; Jia, S. (2013). «Chicken-sized oviraptorid dinosaurs from central China and their ontogenetic implications» (PDF). Naturwissenschaften. 100 (2): 165−175. Bibcode:2013NW....100..165L. PMID 23314810. doi:10.1007/s00114-012-1007-0
- ↑ a b Clark, J. M.; Norell, M. A.; Chiappe, L. M. (1999). «An oviraptorid skeleton from the Late Cretaceous of Ukhaa Tolgod, Mongolia, preserved in an avianlike brooding position over an oviraptorid nest». American Museum Novitates (3265): 1−36. hdl:2246/3102
- ↑ Funston, G. F.; Tsogtbaatar, C.; Tsogtbaatar, K.; Kobayashi, Y.; Sullivan, C.; Currie, P. J. (2020). «A new two-fingered dinosaur sheds light on the radiation of Oviraptorosauria». Royal Society Open Science. 7 (10). 201184 páginas. Bibcode:2020RSOS....701184F. PMC 7657903. PMID 33204472. doi:10.1098/rsos.201184
- ↑ Barsbold, R. (1977). «Kinetism and peculiarity of the jaw apparatus of oviraptors (Theropoda, Saurischia)». Soviet-Mongolian Paleontological Expedition, Trudy. 4: 37−47 Papel traduzido
- ↑ Smith, D. K. (1990). «Osteology of Oviraptor philoceratops, a possible herbivorous theropod from the Upper Cretaceous of Mongolia». Journal of Vertebrate Paleontology. 10 (supp. 003): 42A. doi:10.1080/02724634.1990.10011841
- ↑ Norell, M. A.; Gaffney, E. S.; Dingus, L. (1995). Discovering Dinosaurs In the American Museum of Natural History (em inglês). [S.l.]: Knopf Inc. p. 225
- ↑ Jansen, S. O. K. (2008). Beak morphology in oviraptorids, based on extant birds and turtles (Tese) (em inglês). University of Oslo. pp. 1−48. hdl:10852/11785
- ↑ Funston, G. F.; Mendonca, S. E.; Currie, P. J.; Barsbold, R.; Barsbold, R. (2018). «Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin». Palaeogeography, Palaeoclimatology, Palaeoecology (em inglês). 494: 101−120. Bibcode:2018PPP...494..101F. doi:10.1016/j.palaeo.2017.10.023
- ↑ Hopp, T. P.; Orsen, M. J. (2004). «Dinosaur Brooding Behavior and the Origin of Flight Feathers» (PDF). In: Currie, P. J.; Koppelhus, E. B.; Shugar, M. A.; Wright, J. L. Feathered dragons: studies on the transition from dinosaurs to birds. Bloomington: Indiana University Press. pp. 234−250
- ↑ Sato, T.; Cheng, Y.-N.; Wu, X.-C.; Zelenitsky, D. K.; Hsiao, Y.-F. (2005). «A Pair of Shelled Eggs Inside A Female Dinosaur» (PDF). Science. 308 (5720). 375 páginas. PMID 15831749. doi:10.1126/science.1110578
- ↑ Varricchio, D. J.; Moore, J. R.; Erickson, G. M.; Norell, M. A.; Jackson, F. D.; Borkowski, J. J. (2008). «Avian paternal care had dinosaur origin». Science. 322 (5909): 1826−1828. Bibcode:2008Sci...322.1826V. PMID 19095938. doi:10.1126/science.1163245
- ↑ Yang, T.-R.; Chen, Y.-H.; Wiemann, J.; Spiering, B.; Sander, P. M. (2018). «Fossil eggshell cuticle elucidates dinosaur nesting ecology». PeerJ. 6: e5144. PMC 6037156. PMID 30002976. doi:10.7717/peerj.5144
- ↑ Yang, T.-R.; Wiemann, J.; Xu, L.; Cheng, Y.-N.; Wu, X.-C.; Sander, P. M. (2019). «Reconstruction of oviraptorid clutches illuminates their unique nesting biology». Acta Palaeontologica Polonica. 466: 581−596. doi:10.4202/app.00497.2018
- ↑ a b Dashzeveg, D.; Dingus, L.; Loope, D. B.; Swisher III, C. C.; Dulam, T.; Sweeney, M. R. (2005). «New Stratigraphic Subdivision, Depositional Environment, and Age Estimate for the Upper Cretaceous Djadokhta Formation, Southern Ulan Nur Basin, Mongolia» (PDF). American Museum Novitates (3498): 1−31. doi:10.1206/0003-0082(2005)498[0001:NSSDEA]2.0.CO;2. hdl:2246/5667
- ↑ Jerzykiewicz, T. (1997). «Djadokhta Formation». In: Currie, P. J.; Padian, K. Encyclopedia of Dinosaurs. San Diego: Academic Press. pp. 188−191. ISBN 978-0-12-226810-6
- ↑ a b Chinzorig, T.; Kobayashi, Y.; Tsogtbaatar, K.; Currie, P. J.; Watabe, M.; Barsbold, R. (2017). «First Ornithomimid (Theropoda, Ornithomimosauria) from the Upper Cretaceous Djadokhta Formation of Tögrögiin Shiree, Mongolia». Scientific Reports. 7 (5835): 5835. Bibcode:2017NatSR...7.5835C. PMC 5517598. PMID 28724887. doi:10.1038/s41598-017-05272-6
- ↑ a b Dingus, L.; Loope, D. B.; Dashzeveg, D.; Swisher III, C. C.; Minjin, C.; Novacek, M. J.; Norell, M. A. (2008). «The Geology of Ukhaa Tolgod (Djadokhta Formation, Upper Cretaceous, Nemegt Basin, Mongolia)» (PDF). American Museum Novitates (3616): 1−40. doi:10.1206/442.1. hdl:2246/5916 [ligação inativa]
- ↑ Cau, A.; Beyrand, V.; Voeten, D. F. A. E.; Fernandez, V.; Tafforeau, P.; Stein, K.; Barsbold, R.; Tsogtbaatar, K.; Currie, P. J.; Godefroit, P. (2017). «Synchrotron scanning reveals amphibious ecomorphology in a new clade of bird-like dinosaurs». Nature. 552 (7685): 395−399. Bibcode:2017Natur.552..395C. PMID 29211712. doi:10.1038/nature24679
- ↑ Alicea, J.; Loewen, M. (2013). «New Minotaurasaurus material from the Djodokta Formation establishes new taxonomic and stratigraphic criteria for the taxon». Journal of Vertebrate Paleontology. Program and Abstracts. 76 páginas. Consultado em 19 de novembro de 2020. Arquivado do original em 1 de novembro de 2020

Ligações externas
 Media relacionados com Oviraptor no Wikimedia Commons
Media relacionados com Oviraptor no Wikimedia Commons O Wikispecies possui informações sobre: Oviraptor
O Wikispecies possui informações sobre: Oviraptor
| |||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||


















| Controle de autoridade |
|
|---|
 Portal dos dinossauros
Portal dos dinossauros Portal de anfíbios e répteis
Portal de anfíbios e répteis Portal da paleontologia
Portal da paleontologia Portal da Ásia
Portal da Ásia














