Estoma
No debe confundirse con Estroma.
Para otros usos de este término, véase Estoma (medicina).



En Botánica, se denomina estoma[1] a las células oclusivas que forman parte de la epidermis de la planta y que delimitan entre ellas un poro llamado ostíolo; la epidermis es impermeable al agua y gases debido a la capa de cutícula que la cubre, que permiten comunicar el ambiente gaseoso del interior de la planta con el del exterior. Estas células junto con las células acompañantes de la epidermis alrededor de ellas, si las hay, forman el órgano llamado aparato estomático, que normalmente se abre a una cavidad debajo de la epidermis llamada cámara subestomática cuya función es alojar los gases. Las células oclusivas abren y cierran el ostíolo de forma acoplada con las condiciones fisiológicas de la planta, lo que diferencia al estoma de un poro común como el que puede encontrarse en los musgos más primitivos de las embriofitas (plantas terrestres).
Función
Están localizados en todas las partes herbáceas ("verdes") de las plantas vasculares y algunos musgos (es decir, en la mayoría de las plantas terrestres) que son las partes cuyo tejido de protección es la epidermis, sin crecimiento secundario. Junto con otros órganos de función similar (como las lenticelas, zonas con aspecto verrugoso en la corteza de las partes leñosas cuyas células están separadas por grandes espacios intercelulares, por lo que funcionalmente son poros), son la vía por la que difunde la mayor parte del oxígeno y del dióxido de carbono, dos gases utilizados por las células del interior de la planta durante la fotosíntesis y la respiración celular, y también son la vía principal por la que la planta pierde el agua absorbida por las raíces en forma de vapor de agua (proceso llamado transpiración vegetal). Como el intercambio de gases es positivo pero la pérdida de agua en esa proporción es indeseable, la evolución desarrolló dos estrategias que limitan la pérdida de agua a través de los poros. Una es su ubicación preferentemente en el envés de las hojas, al abrigo del sol directo, y la otra es la evolución de los "estomas" con la posibilidad de abrir o cerrar los poros según las condiciones fisiológicas de la planta.
Ganancia de CO2 y pérdida de agua
El dióxido de carbono, un reactivo clave en la fotosíntesis, está presente en la atmósfera en una concentración de aproximadamente 400 ppm. La mayoría de las plantas requieren que los estomas estén abiertos durante el día. Los espacios de aire de la hoja están saturados de vapor de agua, que sale de la hoja a través de los estomas en un proceso conocido como transpiración. Por lo tanto, las plantas no pueden ganar dióxido de carbono sin perder simultáneamente vapor de agua.[2]
Aproximaciones alternativas
Normalmente, el dióxido de carbono se fija a la ribulosa 1,5-bisfosfato (RuBP) mediante la enzima RuBisCO en las células del mesófilo expuestas directamente a los espacios de aire dentro de la hoja. Esto exacerba el problema de la transpiración por dos razones: en primer lugar, RuBisCo tiene una afinidad relativamente baja por el dióxido de carbono y, en segundo lugar, fija oxígeno en RuBP, desperdiciando energía y carbono en un proceso llamado fotorrespiración. Por ambas razones, RuBisCo necesita altas concentraciones de dióxido de carbono, lo que significa amplias aperturas estomáticas y, como consecuencia, una alta pérdida de agua.
Se pueden utilizar aberturas estomáticas más estrechas junto con una molécula intermediaria con una alta afinidad por el dióxido de carbono, la enzima fosfoenolpiruvato carboxilasa (PEPcase). Sin embargo, recuperar los productos de la fijación de carbono de PEPCase es un proceso que consume mucha energía. Como resultado, la alternativa PEPCase es preferible sólo cuando el agua es limitante pero la luz es abundante, o donde las altas temperaturas aumentan la solubilidad del oxígeno en relación con la del dióxido de carbono, magnificando el problema de oxigenación de RuBisCo.
Plantas con metabolismo ácido de las crasuláceas

Un grupo de plantas en su mayoría desérticas llamadas "CAM" (Crassulacean acid metabolism), las plantas con metabolismo ácido de las crasuláceas, por la familia Crassulaceae, que incluye las especies en las que se descubrió por primera vez el proceso CAM) abren sus estomas por la noche (cuando el agua se evapora más lentamente de las hojas para un determinado grado de apertura de los estomas), usan PEPcase para fijar dióxido de carbono y almacenar los productos en grandes vacuolas. Al día siguiente, cierran sus estomas y liberan el dióxido de carbono fijado la noche anterior en presencia de RuBisCO. Esto satura RuBisCO con dióxido de carbono, permitiendo una fotorrespiración mínima. Sin embargo, este enfoque está muy limitado por la capacidad de almacenar carbono fijo en las vacuolas, por lo que es un sistema preferible sólo cuando la presencia de agua es muy limitada.
Anatomía
Anatómicamente, es decir, a nivel celular al microscopio, se observa que en todos los estomas son dos las células que cambian su tamaño regulando el tamaño del poro, que, como en la figura, lo delimitan, uniéndose entre sí en sus dos extremos. Estas células fueron llamadas "células de guarda" u oclusivas y el poro "ostíolo" (en inglés stoma, mismo nombre que todo el órgano). Por una señal fisiológica, las células oclusivas pueden perder su rigidez de forma que al quedar fláccidas las paredes alrededor del poro se acercan, cerrándolo, y por la señal fisiológica contraria aumentan su turgencia (se hinchan hasta topar con la forma de sus paredes celulares), lo cual expande la célula a lo largo, de forma que se curva alrededor del poro, abriéndolo. La forma en que se curvan las células oclusivas al aumentar su turgencia está decidida por la morfología de sus paredes celulares, la cual puede variar entre taxones. Fisiológicamente, las células de guarda regularían su turgencia mediante un control de la cantidad de sales potasio (K+) en la célula, en relación con la cantidad de sales fuera de ella, de forma que el agua atraviesa la membrana celular hacia adentro o afuera de la célula por ósmosis, es decir, en busca de la mayor concentración de solutos; situación regulada por estructuras presentes en la membrana celular que transfieren sales y otros iones activamente con gasto de energía, lo cual se llama transporte activo y es en esa regulación iónica en la que ayudarían células acompañantes, si las hay. Las células acompañantes, o subsidiarias o anexas, pueden o no estar presentes, y son células epidérmicas contiguas a las de guarda, con una morfología que las distingue del resto de la epidermis.
Todas las plantas terrestres están cubiertas por una capa cerosa secretada por la epidermis que las vuelve casi impermeables (impide la pérdida de agua), llamada cutícula. La cutícula puede ser más gruesa, como en plantas de ambientes xerofíticos, o más delgada, como en plantas de ambientes húmedos, el grosor también puede depender de las condiciones ambientales en que creció la planta y puede ser diferente en diferentes órganos verdes de la planta, pero solo está interrumpida en los ostíolos de los estomas. Esta capa de ceras, no presente en las algas acuáticas de las que se originaron, fue necesaria para su transición a la vida terrestre, ya que impide la extrema desecación que sufrirían si no la tuvieran,[3] pero también impide a la planta captar y liberar los gases que utiliza o desecha en la fotosíntesis y respiración, por lo que paralelamente a la capa de ceras, evolucionaron los diferentes tipos de poros en las plantas, como también el ambiente interno a ellos que aloja una concentración de gases que permite las actividades químicas de la planta. Los primeros poros que aparecieron en las plantas terrestres eran de morfología sencilla y todavía se ven estos "poros" de morfología sencilla en algunos musgos, que pueden tener una cutícula delgada y no están tan adaptados a resistir la desecación. Al originarse las plantas vasculares o traqueofitas (grupo que comprende los helechos y las plantas con semilla), o quizás un poco antes al originarse los musgos hermanos a ellas, la capa de ceras se volvió más gruesa y se originaron los "estomas" de morfología más compleja. (Judd et al. 2007[4])
Los estomas son uno de los participantes en la fotosíntesis y también en la respiración de las plantas, ya que por ellos transcurre el intercambio gaseoso mecánico, es decir que en este lugar sale el oxígeno y entra dióxido de carbono (en la fotosíntesis) y viceversa (durante la respiración) según qué procesos químicos ocurran en las plantas.
La suberina se sintetiza solamente en órganos que desarrollan corcho (felema) como raíces, tallos y algunos frutos.
Criptas estomáticas
Las criptas estomáticas son áreas hundidas de la epidermis de la hoja que forman una estructura similar a una cámara que contiene uno o más estomas y, a veces, tricomas o acumulaciones de cera. Las criptas estomáticas pueden ser una adaptación a la sequía y a las condiciones climáticas secas cuando las criptas estomáticas son muy pronunciadas. Sin embargo, los climas secos no son los únicos lugares donde se pueden encontrar. Las siguientes plantas son ejemplos de especies con criptas o antecámaras estomáticas: Nerium oleander, coníferas, Hakea[5] y Drimys winteri que es una especie de planta que se encuentra en el bosque nuboso.[6]
Los estomas como vías de patogénos
Los estomas son agujeros en la hoja por los que los patógenos pueden entrar sin obstáculos. Sin embargo, los estomas pueden detectar la presencia de algunos, si no de todos, los patógenos.[7] Sin embargo, aplicando bacterias patógenas a las hojas de la planta Arabidopsis estas pueden liberar la sustancia química coronatina,[8] que induce la reapertura de los estomas.[9]
Fisiología de los estomas
La apertura o cierre de los estomas forma parte de la fisiología de la planta que comunica la planta con las condiciones ambientales y la adaptan a ella.
La apertura o cierre de los estomas está muy finamente regulada en la planta por factores ambientales como la luz, la concentración de dióxido de carbono o la disponibilidad de agua. Según investigaciones, se conoce que algunos cationes como el potasio y calcio y aniones como el cloruro intervienen activamente en la apertura y cierre de los estomas. En casos de sequía (estrés hídrico) se cierran los estomas impidiendo pérdidas de agua en la planta, lo cual, sin embargo, también imposibilita el intercambio de gases y, en consecuencia, la entrada de dióxido de carbono (CO2) atmosférico necesaria para la nutrición de las plantas mediante el proceso de fotosíntesis. Es por ello que en regiones xerófilas, los estomas frecuentemente son pequeños o casi inexistentes, y además, contienen cantidades apreciables de ceras, pelos y tricomas, que dificultan la salida del vapor de agua.
El tamaño de estos varía con el tipo de estoma que se está observando, pero por lo general oscila entre 0,6-3,5 micrómetros.
Desarrollo
[cita requerida] Los estomas comienzan su desarrollo cuando una célula "madre", meristemática, sufre una división asimétrica, dando origen a un meristemoide y a una célula hermana de mayor tamaño. A su vez, el meristemoide puede seguir realizando divisiones asimétricas o bien puede diferenciarse hasta transformarse en una célula de guarda madre. Finalmente, la célula de guarda madre sufre una división simétrica y se diferencia en un par de células de guarda maduras alrededor de un poro estomático.
Las células estomáticas tienen una pared interna elástica y de mayor grosor que la externa. Característica importante en el mecanismo de apertura y cierre de los estomas.
Tipos de estomas según disposición de células
Los taxónomos han observado al microscopio los estomas de muchas especies de plantas y han observado que las especies se agrupan según su "tipo de estoma" en grupos de parentesco que tradicionalmente se han ubicado en la categoría de familia, por lo que el "tipo de estoma" es un carácter de valor taxonómico cuando se quiere averiguar el parentesco de un taxón al nivel de esa categoría taxonómica, y muy probablemente no sirva por ejemplo, para diferenciar una especie de otra muy emparentada, ya que probablemente comparten el mismo "tipo de estoma". Los siguientes tipos de estoma son relevantes para el taxónomo de plantas y sólo con figuras acompañantes que dejen claro qué deben observar al microscopio.
- Anomocítico o ranunculáceo: No posee células anexas, es el más frecuente en dicotiledóneas y también el más antiguo. Por Ranunculaceae.
- Paracítico o rubiáceo: Posee 2 células anexas, dispuestas paralelamente con respecto a las oclusivas. Por Rubiaceae.
- Anisocítico o crucífero: Posee 3 células anexas, 1 más pequeña. Por Cruciferae (=Brassicaceae). También en Solanaceae.
- Tetracítico: Posee 4 células subsidiarias. Común en varias familias de monocotiledóneas como Araceae, Commelinaceae, Musaceae
- Diacítico o cariofiláceo: Posee 2 células anexas perpendiculares a las oclusivas. Pocas familias, Cariofiláceas, Acantáceas.
- Ciclocítico: Posee numerosas células subsidiarias, dispuestas en uno o dos círculos alrededor de las células oclusivas.
- Helicocítico: Posee varias células subsidiarias dispuestas en espiral alrededor de las oclusivas.
En los helechos se distinguen cuatro tipos de estomas diferentes:
- Los estomas hipocíticos tienen dos células protectoras en una capa con solo células epidermis ordinarias, pero con dos células subsidiarias en la superficie exterior de la epidermis, dispuestas paralelas a las células protectoras, con un poro entre ellas, superpuesta a la abertura del estoma.
- Los estomas pericíticos tienen dos células protectoras que están completamente rodeadas por una célula subsidiaria continua (como una rosquilla).
- Los estomas desmocíticos tienen dos células protectoras que están completamente rodeadas por una célula subsidiaria que no ha fusionado sus extremos (como una salchicha).
- Los estomas polocíticos tienen dos células protectoras que están rodeadas en gran medida por una célula subsidiaria, pero que también están en contacto con células epidérmicas ordinarias (como una U o una herradura).
Evolución
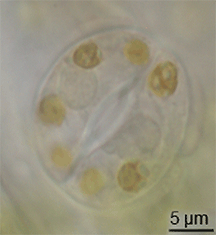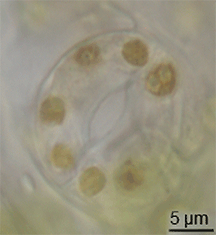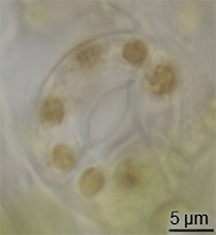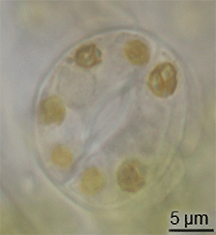
Hay poca evidencia de la evolución de los estomas en el registro fósil, pero habían aparecido en plantas terrestres a mediados del período Silúrico.[10] Es posible que hayan evolucionado mediante la modificación de conceptáculos de ancestros parecidos a las algas de las plantas.[11] Sin embargo, la evolución de los estomas debe haber ocurrido al mismo tiempo que evolucionaba la cutícula cerosa; estos dos rasgos juntos constituyeron una gran ventaja para las primeras plantas terrestres.
Véase también
Referencias
- ↑ J Valla, Morfología de Plantas Superiores
- ↑ Debbie Swarthout and C.Michael Hogan. 2010. Stomata. Encyclopedia of Earth. National Council for Science and the Environment, Washington DC
- ↑ Kenrick P, Crane PR. 1997. The Origin and Early Evolution of Land Plants. Smithsonian Institution Press, Washington, DC.
- ↑ Judd, WS; CS Campbell, EA Kellogg, PF Stevens, MJ Donoghue. 2007. "An overview of green plant phylogeny". En: Plant Systematics, a phylogenetic approach. Third edition. Capítulo 7.
- ↑ Jordan, Gregory J; Weston, Peter H; Carpenter, Raymond J; Dillon, Rebecca A.; Brodribb, Timothy J. (2008). «The evolutionary relations of sunken, covered, and encrypted stomata to dry habitats in Proteaceae». American Journal of Botany 95 (5): 521-530. doi:10.3732/ajb.2007333.
- ↑ Roth-Nebelsick, A.; Hassiotou, F.; Veneklaas, E. J (2009). «Stomatal crypts have small effects on transpiration: A numerical model analysis.». Plant Physiology 151 (4): 2018-2027. PMC 2785996. PMID 19864375. doi:10.1104/pp.109.146969.
- ↑ Maeli Melotto; William Underwood; Jessica Koczan; Kinya Nomura; Sheng Yang He (2006). «Plant Stomata in innate immunity against bacterial invasion». Cell (en inglés) 126 (5): 969-980. PMID 16959575. S2CID 13612107. doi:10.1016/j.cell.2006.06.054.
- ↑ La coronatina (COR) es una toxina producida por la bacteria Pseudomonas syringae. Participa en hacer que los estomas se vuelvan a abrir después de que se cierran en respuesta a patrones moleculares asociados a patógenos, además de interferir con las respuestas mediadas por el ácido salicílico después de que ha comenzado la infección. Consiste en ácido coronafácico (CFA), que es un análogo del ácido metil jasmónico (MeJA), y ácido coronamico (CMA), unidos por un enlace amida entre el grupo ácido del CFA y el grupo amino del CMA (Melotto, Maeli; Underwood, William; He, Sheng Yang (2008). "Role of Stomata in Plant Innate Immunity and Foliar Bacterial Diseases". Annual Review of Phytopathology. 46: 101–22. doi:10.1146/annurev.phyto.121107.104959. PMC 2613263. PMID 18422426 )
- ↑ Schulze-Lefert, P; Robatzek, S (2006). «Plant pathogens trick guard cells into opening the gates». Cell 126 (5): 831-834. doi:10.1016/j.cell.2006.08.020. hdl:11858/00-001M-0000-0012-394E-B.
- ↑ D. Edwards, H. Kerp; Hass, H. (1998). «Stomata in early land plants: an anatomical and ecophysiological approach». Journal of Experimental Botany (en inglés) 49 (Special Issue): 255-278. doi:10.1093/jxb/49.Special_Issue.255.
- ↑ Krassilov, Valentin A. (2004). «Macroevolutionary events and the origin of higher taxa». En Wasser, Solomon P., ed. Evolutionary theory and processes : modern horizons : papers in honour of Eviatar Nevo. Dordrecht: Kluwer Acad. Publ. pp. 265-289. ISBN 978-1-4020-1693-6.
- Gola, G., Negri, G. y Cappeletti, C. 1965. Tratado de Botánica. 2.ª edición. Editorial Labor S.A., Barcelona, 1110 p.
- Font Quer, P. 1953. Diccionario de Botánica. Barcelona, Labor.
- Strassburger, E. 1994. Tratado de Botánica. 8.ª edición. Omega, Barcelona, 1088 p.
- Morfología de Plantas Vasculares. Cátedra de la UNNE. Estomas. https://web.archive.org/web/20090918074833/http://www.biologia.edu.ar/botanica/tema13/13-6clasifest.htm (al 25/09/14)
- Stuart Casson and Julie E. Gray. Influence of environmental factors on stomatal development.. New Phytol. 2008;178(1):9-23
.
Enlaces externos
 Wikimedia Commons alberga una categoría multimedia sobre Estoma.
Wikimedia Commons alberga una categoría multimedia sobre Estoma.- Morfología de Plantas Vasculares. Cátedra de la UNNE. https://web.archive.org/web/20090918074833/http://www.biologia.edu.ar/botanica/tema13/13-6clasifest.htm
 Datos: Q216635
Datos: Q216635- Multimedia: Stoma / Q216635






